
| Bio-Ecke | Chemie-Ecke | Home | Unterricht | sOWLrebel |
|---|
Alle Lipide besitzen ausgedehnte unpolare Molekülteile, sind daher wasserunlöslich und lassen sich aus Zellstrukturen nur mit unpolaren Lösungsmitteln extrahieren.
Befindet sich "am anderen Ende" des Moleküls eine polare Gruppe, so bewirkt dieser amphipathische Bau eine hohe Grenzflächenaktivität. Daraus erklärt sich die Bedeutung dieser Lipide für den Aufbau biologischer Membranen.
Die Hauptgruppe der Lipide bilden die Fette, die als Speicherstoffe in Zellen abgelagert werden. Andere Lipide sind biologisch aktiv: So zählen manche Vitamine bzw. deren Vorstufen und einige Hormone zu dieser Stoffgruppe. Die Lipide werden in Fette (nicht hydrolysierbar) und Lipoide oder zusammengesetzte Lipide (nicht hydrolysierbar, fettähnliche Stoffe) untergliedert.
Fettspeicherung erfolgt im Samen höherer Pflanzen entweder im Endosperm (z.B:Rhizinus) oder in den Cotyledonen (=Keimblätter) (z.B.:Cruciferae). Oft sind die Zellen dann von zahlreichen, (membranumschlossenen?) Fetttropfen erfüllt: Oleosomen, an die sich dann mit Keimungsbeginn viele Glyoxysomen, die spezifischen Organellen des Abbaus, anlagern. Reichlich Fett enthält auch das Fruchtfleisch der Olive, welches zusammen mit einigen anderen Samen und Früchten (Lein, Raps, Sonnenblume, Erdnuss, Baumwolle, Cocosnuss) als Ausgangsmaterial zur technischen Öl-bzw. Fettgewinnung dient. Auch Algen (Grünalgen, Diatomeen) können Fett anhäufen, und zwar bis zu 65% ihrer Trockenmasse.
Die Naturfette sind in der Regel keine Reinstoffe. Sie stellen Gemische aus verschiedenen Glycerinen dar (sog. heterogene Fette). Anstelle eines exakt definierten Schmelzpunktes besitzen sie daher ein Erweichungsintervall.
Von der Weltfettproduktion entfielen 1980 zwei Drittel auf pflanzliche Fette (Kokosfett, Palmkernfett) und fette Öle (Olivenöl, Sojabohnenöl, Sonnenblumenöl, Palmöl) und ein Drittel auf tierische Fette (Butter, Talg, Schweineschmalz, Waltran).
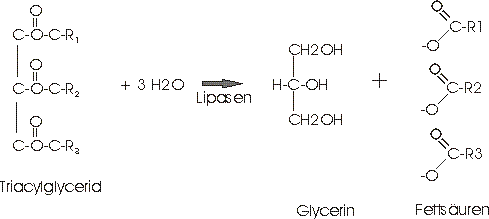
Die Fette lassen sich hydrolytisch spalten. Dabei werden Glycerin und Fettsäuren bzw. Fettsäuresalze gebildet.
Für Butter wurde eine Verseifungszahl von 230, für Erdnußöl von nur 158 gefunden. Erdnußbutter enthält in seinen Fettmolekülen also wesentlich längere Fettsäuren (zu 84% mit C18 und zu 10% mit C16) als Buttersäure (30% mi C16, 46% mit C18, der Rest von C4 bis C14).

Die in Naturfetten enthaltenen Carbonsäuren zeigen mehrere Übereinstimmungen:

Infolge der cis-Konfiguration an der Doppelbindung entsteht ein bedeutsamer Knick in der Molekülkette. Daher erschweren ungesättigte Fettsäuren eine regelmäßige Anordnung der Fettmoleküle im Kristallgitter. Mit zunehmendem Anteil ungesättigter Fettsäuren sinkt die Erweichungstemperatur und die Fette werden (bei Zimmertemperatur) immer weicher. Kokosfett schmilzt zwischen 24° und 27°C, Butterfett zwischen 31° und 36°C, während die "fetten Öle" Olivenöl erst bei -2°C und Sonnenblumenöl bei -18°C fest werden.
Bei dieser katalytischen Hydrierung werden die Öle mit Nickelpulver versetzt und bei 180°C und 6 bar Druck von Wasserstoffgas durchspült.
Linolsäure (C18H32O2) und Linolensäure (C18H30O2) treten gehäuft im Leinöl auf.


Beide Verbindungen gehören auch zu den Nahrungsbestandteilen, die für Mensch und Tier essentiell sind. Sie werden zum Teil umgewandelt in die Eicosapentaensäure, welche unter anderem gegen Herzinfarkt schützen soll.
Während Ölsäure und Palmitinsäure (gesättigt!) in den pflanzlichen Reservefetten weit verbreitet sind, treten andere Fettsäuren gehäuft in speziellen Zellorganellen auf, so z.B. die Linolensäure in den Chloroplasten.
Weitere ungesättigte Fettsäuren finden sich in unterschiedlicher Verteilung und Quantität in den Fetten einzelner Pflanzenarten und -familien.
In den Sekundärcarotinoiden sind Xanthophylle esterartig an gesättigte Fettsäuren gebunden (Bsp: Luteinbispalmitat).
Als Maß für die Konzentration freier Fettsäuren in einem Fett dient die Säurezahl, die angibt, wie viele mg Kaliumhydroxid zur Neutralisation (nicht Verseifung!) von einem Gramm Fett benötigt werden.
Dementsprechend finden wir in den meisten Phospholipiden eine Phosphatidsäure als Grundgerüst. Solche Verbindungen heißen Glycerinphosphatide; zusammen mit den Glykolipiden bilden sie die Gruppe der Glycerolipide. Sie werden den Sphingolipiden gegenübergestellt, bei denen Diacylglycerin durch Sphingosin mit säureamidgebundener Fettsäure ersetzt ist.
Die beiden Membranen pflanzlicher Mitochondrien weisen qualitativ offensichtlich die gleiche Phospholipid Zusammensetzung auf, wobei Phosphatidyl-cholin und -ethanolamin die Hauptbestandteile sind. Die Aussenmembran ähnelt dem Plasmalemma insofern, als sie doppelt so viel Phosphatidyl-cholin als -ethanolamin enthält; in den Innenmembranen sind ihre Mengen gleich. Beide Verbindungen haben dort mehr ungesättigte Fettsäure-Reste als in der Aussenmembran. In den Chloroplasten sind neben den dominierenden Galaktolipiden auch Glycerinphosphatide vorhanden: -cholin tritt gehäuft in der Hülle; -glycerin in deutlich geringeren Mengen in Hülle und Thylakoidmembranen auf.

Solche Lipoide sind in allen Zellmembranen enthalten; besonders reich sind sie in Nervenzellen und Eigelb enthalten. Ein bekannter Vertreter ist das Lecithin (Cholin-Phosphoglycerid). Das Molekül besitzt neben einem stark polaren Kopf zwei unpolare Schwänze und zeigt demnach amphipathische Eigenschaften.

Auch andere Verbindungen dieses Typs können seine Position einnehmen, so bei höheren Pflanzen und Hefen das reduzierte und um eine Hydroxygruppe erweiterte Hydroxydihydrosphingosin.

Das N-Acylsphingosin trägt an ihrer Aminogruppe stets einen Fettsäurerest; diese Säureamide heißen Ceramide.

Aus den Ceramiden gehen sowohl die Sphingomyeline (durch Anlagerung von Cholin) als auch die Ganglioside (über Cerebrosid durch Anlagerung eines Zuckers).
Die höchste Konzentration an Gangliosiden findet man im Nervensystem, speziell in der grauen Substanz, wo sie 6% aller Lipide ausmachen. Ganglioside werden kontinuierlich abgebaut, und zwar durch sequentielle Abspaltung der endständigen Zucker durch hochspezifische Glykosylhydrolasen. Der Gangliosidabbau findet in den Lysosomen statt. Organellen, die viele spaltende Enzyme enthalten und auf den geordneten Abbau von Zellkomponenten spezialisiert sind.
Störungen des Gangliosidabbaus können ernste klinische Konsequenzen haben. Bei der Tay-Sachs-Krankheit werden die Symptome sichtbar, noch bevor das betroffene Kind ein Jahr alt ist. Schwäche und verzögerte psychomotorische Entwicklung sind typische Frühsymptome; wenige Monate später verschlechtert sich die Sehkraft, und es treten spastische Lähmungen auf. Mit zwei Jahren ist das Kind schwachsinnig und blind. Gewöhnlich endet die Krankheit mit Erreichen des dritten Lebensjahres tödlich.
Auffällige pathologische Veränderungen finden im Nervensystem statt, wo die Neuronen durch lipidgefüllte Lysosomen prall anschwellen. Der Gangliosidgehalt des Gehirns eines Kindes mit Tay-Sachs-Syndrom ist dramatisch erhöht. Ursache dieser hohen Konzentration ist das Fehlen des Enzyms, das für den Abbau sorgt.
Die Krankheit wird autosomal rezessiv vererbt, sie kan schon während der fetalen Entwicklungsphase diagnostiziert werden; dazu entnimmt man durch Amniozentese Amnionflüssigkeit (Fruchtwasser) und untersucht sie auf das abbauende Enzym.
Bei höheren Pflanzen dominiert die Galaktose. Unter den Fettsäure-Bausteinen dominiert die Linolensäure, bei einigen Spezies findet sich zusätzlich Hexadecatriensäure. Die drei folgenden Galaktolipide stellen das größte Kontingent der Chloroplastenlipide: Monogalktosyl-diacylglycerin, Digalaktosyl-diacylglycerin, Trigalaktosyl-diacylglycerin.
Hinzu kommt exklusiv bei Pflanzen Sulfochinovosyl-diacylglycerin (ein Sulfolipid) mit dem Zuckerbaustein 6-Deoxy-6-sulfo-D-Glucose.
Auch für Suberin steht die endgültige Strukturaufklärung noch aus; die Verbindung enthält beträchtliche Mengen an veresterten gamma-Hydroxyfettsäuren, Dicarbonsäuren, relativ langen Säuren und Alkoholen (> C20) sowie kleine Anteile von stärker oxidierten Monomeren, welche auch im Cutin vorkommen.
Wachse als Ester langkettiger unverzweigter Fettsäuren (C26-C36) mit langen aliphatischen (C16-C36) oder cyclischen Alkoholen werden im Wechsel mit Suberin bzw. Cutin schichtweise bei der Verkorkung aufgelagert.

Als biologische Synthesevorstufe dieser Baueinheit ist das Isopentenyldiphosphat - aktiviertes Isopren - erkannt worden.

In Pflanzen leiten sich eine Reihe von Verbindungen vom Isopren ab:
Das Cholesterin kommt in Membranen vor, genauso wichtig sind Steroidhormone und Gallensäure die ebenfalls zu den Steroiden zählt.

Cholesterin ist die Vorstufe der fünf Hauptklassen von Steroidhormonen: für Gestagene, Glucocorticoide, Mineralcorticoide, Androgene und Östrogene.
Progesteron, ein Gestagen, bereitet den Uterus für die Aufnahme des befruchteten Eies vor; es ist ausserdem für die Aufrechterhaltung der Schwangerschaft verantwortlich
Androgene (wie etwa Testosteron) bewirken die Ausprägung der sekundären männlichen, Östrogene (zum Beispiel Östron) die der sekundären weiblichen Geschlechtsmerkmale. Östrogene wirken außerdem am Eireifungszyklus mit.
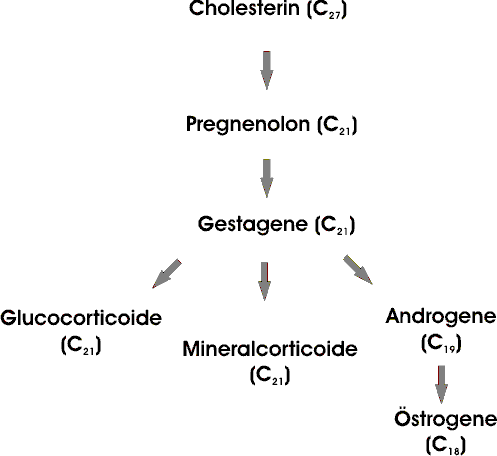
Glucocorticoide (wie Cortisol) fördern die Gluconeogenese und die Glykogenbildung und steigern darüber hinaus den Fett- und Proteinabbau.
Die Mineralcorticoide (hauptsächlich Aldosteron) fördern in den distalen Tubuli der Nieren die Rückresorption von Na+ und die Ausscheidung von K+ und H+ was zum Anstieg von Blutvolumen und Blutdruck führt.
Diese Hormone werden hauptsächlich in folgenden Organen synthetisiert: Gestagene im Corpus luteum (Gehirn), Östrogene in den Ovarien, Androgene in den Hoden, Glucocorticoide und Mineralcorticoide in der Nebennierenrinde.
 Ebenfalls zu den Isoprenderivaten gehören die fettlöslichen Vitaminen.
Ebenfalls zu den Isoprenderivaten gehören die fettlöslichen Vitaminen.Als Vitamine werden alle Spurensubstanzen bezeichnet, die für eine normale Zellfunktion nötig sind, dabei aber vom jeweiligen Organismus nicht selbst synthetisiert werden können. So ist das in Pflanzen verbreitete beta-Carotin als Provitamin unentbehrlicher Bestandteil unserer Nahrung.
 Durch eine oxidative Spaltung genau in der Molekülmitte werden daraus zwei Moleküle Vitamin A gebildet.
Durch eine oxidative Spaltung genau in der Molekülmitte werden daraus zwei Moleküle Vitamin A gebildet.
Die aus 40 Kohlenstoffatomen bestehende Grundskelett der Carotinoide setzt sich aus 8 Baueinheiten des Isoprens zusammen. Diese Einheiten sind so angeordnet, dass eine kettenförmige zentrale Struktur mit 14 Kohlenstoffatomen und 7 konjugierten Doppelbindungen sowie 4 Methyl-Gruppen als Seitenketten entsteht. Sie trägt eine gruppierung an einem oder an beiden Enden, die als offene oder geschlossene Ringstruktur ausgebildet ist; liegen zwei endständige Ringe vor, können sie auch unterschiedlich sein.
Durch die lineare Anordnung der Isopren-Einheiten entsteht ein relativ langes System konjugierter Doppelbindungen. Dieser Molekülaufbau bedingt, dass alle Carotinoide lebhaft gefärbte Verbindungen sind. Sie absorbieren vor allem blaue und violette Anteile der sichtbaren Strahlung und unterstützen den Photosyntheseapparat der Pflanzen.
Die an der Photosynthese beteiligten Carotinoide werden als "Primärcarotinoide" bezeichnet, und den "Sekundärcarotinoiden" gegenübergestellt, die in vielfältiger Form vor allem in Blüten und Früchten als Bestandteil von Chromoplasten, aber auch in heterotrophen Organismen wie Bakterien, Hefen und Pilzen vorkommen. Sekundärcarotinoide entstehen auch in photosynthetisch aktiven Organismen als Folge unzureichender Mineralsalz-Ernährung.
Für die Isolierung von Carotinoiden wird die Säulenchromatographie und die Dünnschichtchromatographie benutzt.

In Wasser entstehen Doppelschichten mit den hydrophoben Molekülteilen im Inneren und den polaren Gruppen an den Grenzflächen zum Wasser. Dies ist im Hinblick auf die natürlichen Zellmembranen von Wichtigkeit.
Biologische Membranen bestehen im Mittel zu 40% aus Lipiden und zu 60% aus Proteinen. Sie sind für Wasser und unpolare Moleküle gut, für polare Moleküle wie Zucker weniger und für anorganische Ionen kaum durchlässig.
2) Das Glycerin geht als Dihydroxyacetonphosphat,in die Glykolyse ein, um dort weiter verwertet zu werden:
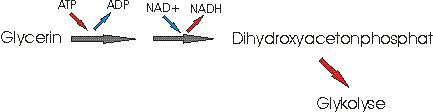
Hierbei wird 1 ATP verbraucht und ein NADH gewonnen, welches in der Atmungskette in 3 ATP umgewandelt wird.
3) Die Fettsäuren werden, unter ATP Verbrauch, mit dem Coenzym A verbunden und zur weiteren Verwertung, als Acyl-CoA in die Mitochondrien geschleust.
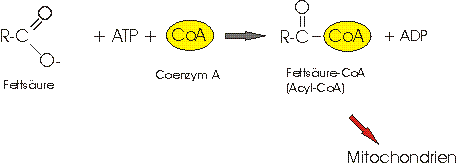
4) Im Mitochondrium geht die Umsetzung weiter. Die Acyl-CoA Kette wird in mehreren Schritten sukzessiv in kleine Acetyl-CoA-Einheiten gespalten.
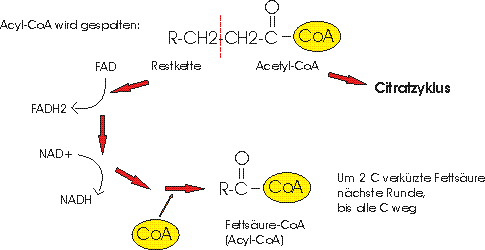
Das Acetyl-CoA wird dann im Citratzyklus verstoffwechselt und liefert dort die üblichen Mengen an NADH, FADH und GTP (ATP).